首页
业务介绍
▼
SCI评估
SCI编译
实验外包
数据统计
课题申请辅导
信誉保障
SCI期刊
联系我们
Science子刊:Piezo1通道在成纤维细胞中起关键作用
时间 : 2024-10-27
1. 促纤维化巨噬细胞与成纤维细胞急性接触可依赖巨噬细胞αvβ3整合素和成纤维细胞Piezo1激活成纤维细胞。
2. Piezo1通道在成纤维细胞中起着核心作用,它能够响应巨噬细胞的接触信号,引发细胞内钙离子水平的变化,促进成纤维细胞的活化。
近日,
“Science Advances”(IF=11.7)
上发表了一篇题为
“Acute contact with profibrotic macrophages mechanically activates fibroblasts via αvβ3 integrin–mediated engagement of Piezo1”
的文章。这篇文章
研究了在纤维化过程中,促纤维化巨噬细胞如何通过机械方式激活成纤维细胞,以及这一过程中涉及的分子机制。
研究背景介绍
纤维化
是一种病理过程,其中正常的组织被异常的纤维组织所替代,导致器官结构和功能的破坏。纤维化可发生在多个器官,如肺、肝和肾,是许多疾病进展的关键因素。
肌成纤维细胞
是一种具有收缩能力的细胞类型,它们在组织修复和纤维化过程中起着核心作用。
Piezo1
是一种机械敏感离子通道,它在细胞对机械刺激的感知中起重要作用。
研究思路分析
研究技术路线图
0
1
巨噬细胞与成纤维细胞的相互作用及其信号通路
①
研究提出,巨噬细胞(Mϕ)通过与成纤维细胞初次接触,通过启动TGF-β1信号或机械刺激(或两者兼有)来激活成纤维细胞。研究中,通过用巨噬细胞集落刺激因子(M-CSF)(=M
M-CSF
)处理培养的小鼠骨髓来源的单核细胞,再分别用
LPS(=M
LPS
,促炎
)
或
IL-4加IL-13(=M
IL-4/13
,促纤维化
)
处理来极化初级Mϕ。
②
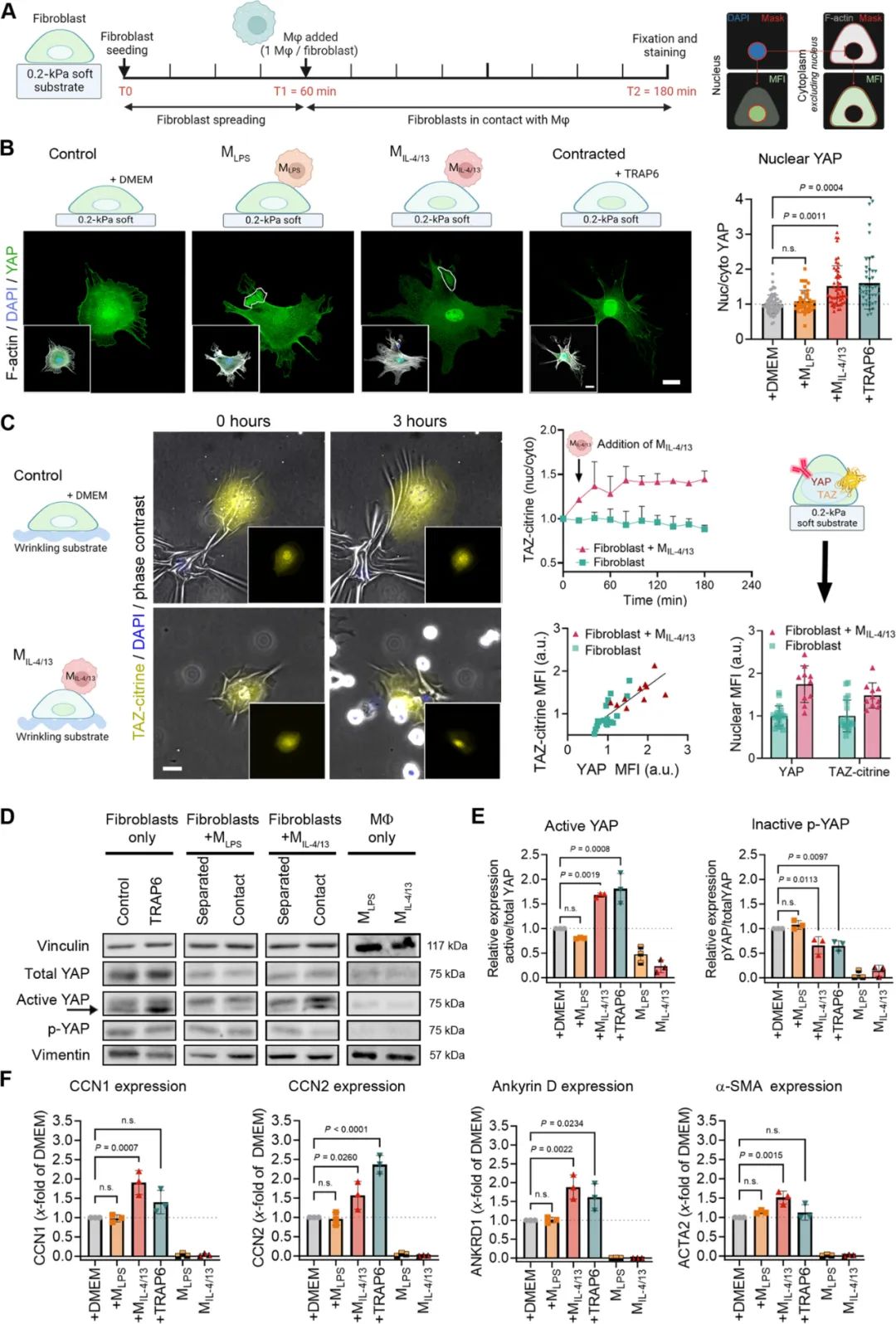
追踪
转录因子YAP
和
转录共激活因子TAZ
,作为机械肌成纤维细胞激活的指标。实验结果表明,与
M
IL-4/13
接触
可以迅速通过YAP/TAZ信号途径
激活成纤维细胞
,而M
LPS
则没有这种效果。此外,在
软质基质上
,M
IL-4/13
并不通过Smad2/3信号通路
激活成纤维细胞,而在
硬质基质
上则可以观察到Smad2/3的核转位。表明,巨噬细胞的
极化状态
和
基质的机械性质
共同影响成纤维细胞的激活和转化。
③
研究发现,
M
IL-4/13
接触活的成纤维细胞时,TAZ-citrine核易位且弹性硅胶底物变形增强,推测其可启动ECM与肌成纤维细胞激活的正反馈回路。实验使用了荧光标记弹性收缩表面(FLECSs)来量化收缩程度。结果显示,
与M
IL-4/13
接触可提高成纤维细胞收缩率和速度
,而M
LPS
无显著影响。
④
此外,增加M
IL-4/13
与成纤维细胞的
接触数量可以进一步增强
收缩反应,而M
LPS
则没有这种效应。还使用三维胶原蛋白凝胶来观察成纤维细胞的局部收缩反应,发现M
IL-4/13
接触可以在
局部区域显著提高成纤维细胞的收缩能力
。表明,促纤维化巨噬细胞通过特定的机械信号传导途径激活成纤维细胞,促进了纤维化过程中的细胞收缩和基质硬化。
0
2
局部力学变化与细胞收缩
①
通过将粒子图像测速技术(PIV)转换为牵引力显微图像,发现与M
IL-4/13
接触后,成纤维细胞的
局部收缩应力显著增加
。此外,在硬质玻璃基质上培养成纤维细胞,发现M
IL-4/13
接触
增强了成纤维细胞中磷酸化肌球蛋白轻链(p-MLC)的染色强度
,这是肌球蛋白激活的一个指标。M
IL-4/13
接触还导致成纤维细胞在接触区域发生
局部膜回缩
,减少了细胞周长的“实体性”和“圆形度”。还发现在与M
IL-4/13
接触区域的
膜张力和局部硬度较高
,这与p-MLC的积累一致。表明,与M
IL-4/13
的接触而非M
LPS
的接触,会在接触点引发成纤维细胞的收缩和机械应力。
②
p-MLC由肌球蛋白轻链激酶(MLCK)调控,MLCK受胞质Ca
2+
浓度调控,通过转染GCaMP6s,一种在结合Ca
2+
时发光的蛋白质,观察到与M
IL-4/13
接触后5分钟内
荧光增加
,表明Ca
2+
浓度升高。此外,
核因子活化T细胞1(NFAT1)
的水平在接触M
IL-4/13
的成纤维细胞中
比未接触的细胞高1.7倍
,进一步证实了Ca
2+
的激活。表明,与M
IL-4/13
的接触可以引发成纤维细胞内Ca
2+
浓度的增加,并导致NFAT1的核转位,而与M
LPS
的接触则没有这种效果。
0
3
分子机制与药物干预
①
测试了两种可能的物理接触依赖性情景:(i)通过异细胞间隙连接直接从Mϕ向成纤维细胞传递钙离子;(ii)巨噬细胞触摸导致成纤维细胞膜上的拉伸激活的离子通道(SACs)开放。结果显示,虽然成纤维细胞和M
IL-4/13
都表达Cx43蛋白,但它们似乎没有形成功能性间隙连接来介导小分子(如钙离子)的直接交换。进一步的研究表明,
SACs对于M
IL-4/13
诱导的细胞内钙离子升高
以及随后的成纤维细胞
收缩是必需的
。特别是
Piezo1通道
,在感应M
IL-4/13
接触方面发挥关键作用,介导细胞内钙离子增加,从而驱动成纤维细胞的收缩。
②
通过筛选单细胞RNA测序数据,发现
整合素β3亚单位(CD61)
在M
IL-4/13
中独特表达,而在M
LPS
中几乎不表达。进一步研究发现
αv和β3整合素
在大多数M
IL-4/13
表面表达,但在M
LPS
中仅少量表达。用
αvβ3整合素抑制剂cyclo(-RGDfK)和β3整合素阻断抗体Axum4
处理M
IL-4/13
,发现这些抑制剂
减少了
接触诱导的Ca
2+
依赖的NFAT1
核转位和成纤维细胞收缩
。
③
在博来霉素诱导的
肺纤维化小鼠模型
中,证实了
β3整合素在促纤维化巨噬细胞中的表达
,并在高度纤维化的肺组织中与α-SMA阳性的肌成纤维细胞相邻。表明M
IL-4/13
上αvβ3整合素的表达对于促进急性接触介导的成纤维细胞激活是必需的。
结论与讨论
本研究发现
M
IL-4/13
通过其表面的αvβ3整合素
与成纤维细胞接触,迅速激活成纤维细胞内的
Piezo1通道,
引发细胞内
钙离子浓度升高和YAP/TAZ信号通路激活
,从而导致成纤维细胞转化为肌成纤维细胞,促进纤维化过程。
未来的研究需要进一步探索αvβ3整合素与成纤维细胞膜上潜在配体的相互作用,以及这些相互作用如何调节Piezo1通道的开启和细胞内钙离子的变化。此外,还需要验证Piezo1通道激活与YAP/TAZ信号通路之间的直接联系,以及NFAT1在肌成纤维细胞激活中的作用。这些研究将有助于深入理解纤维化的分子机制,并为开发新的治疗策略提供科学依据。